卡哈尔体
发现历程与命名由来
1903年,西班牙病理学家Santiago Ramón y Cajal在采用硝酸银染色法对神经元进行观察时,于细胞核内发现了一些与核仁相连接的球状结构,最初将其称为“核仁辅助体”。在随后的几十年里,这一结构在多个物种中被反复观察到,由于不同领域研究者的视角差异,它曾先后被命名为球状细胞器、内体、核仁体及螺旋体等多种称谓。 ADSFAEQWER353423413434
1969年,A. Monneron和W. Bernhard在哺乳动物肝细胞间期核中重新发现了这些细胞器,并基于它们在电镜图像中呈现出螺旋状细丝缠绕的形态特征,将其命名为“螺旋体”。为了避免命名混乱并纪念Cajal的原始发现,1999年Gall等人提议将其统一命名为卡哈尔体。1999年国际学术界正式将这一结构命名为“Cajal body”,中文译名确定为“卡哈尔体”。 ADSFAEQWER353423413434
结构特征
形态学表现
卡哈尔体在电镜下呈现为球形小体样结构,具有无膜细胞器(也称为生物分子凝聚体)的典型特征——其三维结构并非由脂质双分子层划定边界,而是通过内部特定成分(主要为Coilin蛋白)的相互作用,在核质中自发地凝聚形成。它的直径通常在0.1至2.0微米之间,在神经元等特定细胞类型中可达更大尺寸。在同一细胞的细胞核内,典型数目为1至6个,在不同物种、组织和细胞分化阶段中可发生变化。 ADFASDFAF23RQ23R
动态可塑性
卡哈尔体最引人注目的特征之一是它的高度动态性。在细胞分裂的M期,卡哈尔体发生解组装并随细胞质分配;当细胞进入G1/S期时,在活跃的转录和生长信号的驱动下重新组装,在G1/S期达到最大。多个卡哈尔体可以相互靠近并融合成一个,单个卡哈尔体也可能分裂成两个。此外,其组成蛋白和RNA与周围的核浆物质持续发生交换,维持着一种动态平衡状态。卡哈尔体在静止期细胞中消失,在增殖期(如干细胞、肿瘤细胞)中完成重组装。
ADFASDFAF23RQ23R
分子组成
卡哈尔体的分子组成十分复杂。已知的组成蛋白有几十种,核心蛋白包括p80 Coilin(结构支架)、NOPP140(RNA结合蛋白)以及纤维蛋白复合体。
ADFASDFAF23RQ23R
- p80 Coilin蛋白:Coilin是卡哈尔体的标志物和关键支架蛋白,在后生动物中进化保守。它通过与其他蛋白相互作用,提高其支架区域内多种分子的浓度和反应效率。Coilin含有对称性二甲基化的精氨酸,其磷酸化状态调节着卡哈尔体的组装。
此外,卡哈尔体中还富集参与mRNA剪接的各种小核糖核蛋白的组分、snoRNAs、scaRNAs以及其他相关蛋白。2017年发表于Developmental Cell的研究显示,Fam118B蛋白同样作为卡哈尔体的一个重要组分,是其形成、snRNP生物发生和细胞活力所必需的。一些与细胞增殖和细胞周期功能相关的功能分子,也被发现存在于卡哈尔体之中或周边。
ADSFAEQWER353423413434
形成机制:液-液相分离与SMN复合体的级联调控
近年来,卡哈尔体的形成机制得到了深入研究。研究人员发现,Coilin蛋白是通过一种被称为“液-液相分离”的机制在细胞核内发生凝聚,形成了卡哈尔体。这一过程受到SMN复合体蛋白的级联调控——SMN复合体在细胞质中协助snRNP的初步组装;组装完成后,snRNP和SMN一同迁移至细胞核内,与卡哈尔体中的Coillin蛋白相互作用,在Coilin的“包围”下逐步完成最终成熟过程。 ADFASDFAF23RQ23R
2024年,一项发表于Journal of Cell Science的研究进一步揭示了SMN与Coilin之间的空间分离机制。研究人员利用超分辨显微镜和β-咔啉生物碱harmine(一种可逆破坏卡哈尔体的工具化合物)发现:将harmine洗脱后,SMN和Coilin首先形成小的互连凝聚体,SMN凝集体在Coilin的包围下发育为球状结构,最终分离成不同的凝聚体。这一发现强调了SMN多聚化在促进其从Coilin中分离、确保持续snRNP生物发生中的重要性。上海科技大学生命学院的Tomoyasu Sugiyama课题组在2024年也证实,裂殖酵母中的Coilin同源物Mug174能够通过相分离机制形成卡哈尔体,并揭示其维持细胞静息期(G0期)的调控机制,这为理解卡哈尔体功能障碍与人类疾病之间的联系提供了新的视角。 ADSFAEQWER353423413434
功能机制
snRNP生物发生与剪接体组装
卡哈尔体的主要功能之一是作为snRNP(小核核糖核蛋白)成熟的场所,这些snRNP最终构成RNA剪接体。新转录的snRNA被转运到细胞质中发生3′端修饰、5′帽结构超甲基化和SM蛋白结合,然后重新进入细胞核并进入卡哈尔体,在此结合snRNP特异性蛋白、完成剪接体组装所需的关键步骤。核小核糖核蛋白在卡哈尔体中完成2′-O-甲基化和假尿苷化等修饰过程。
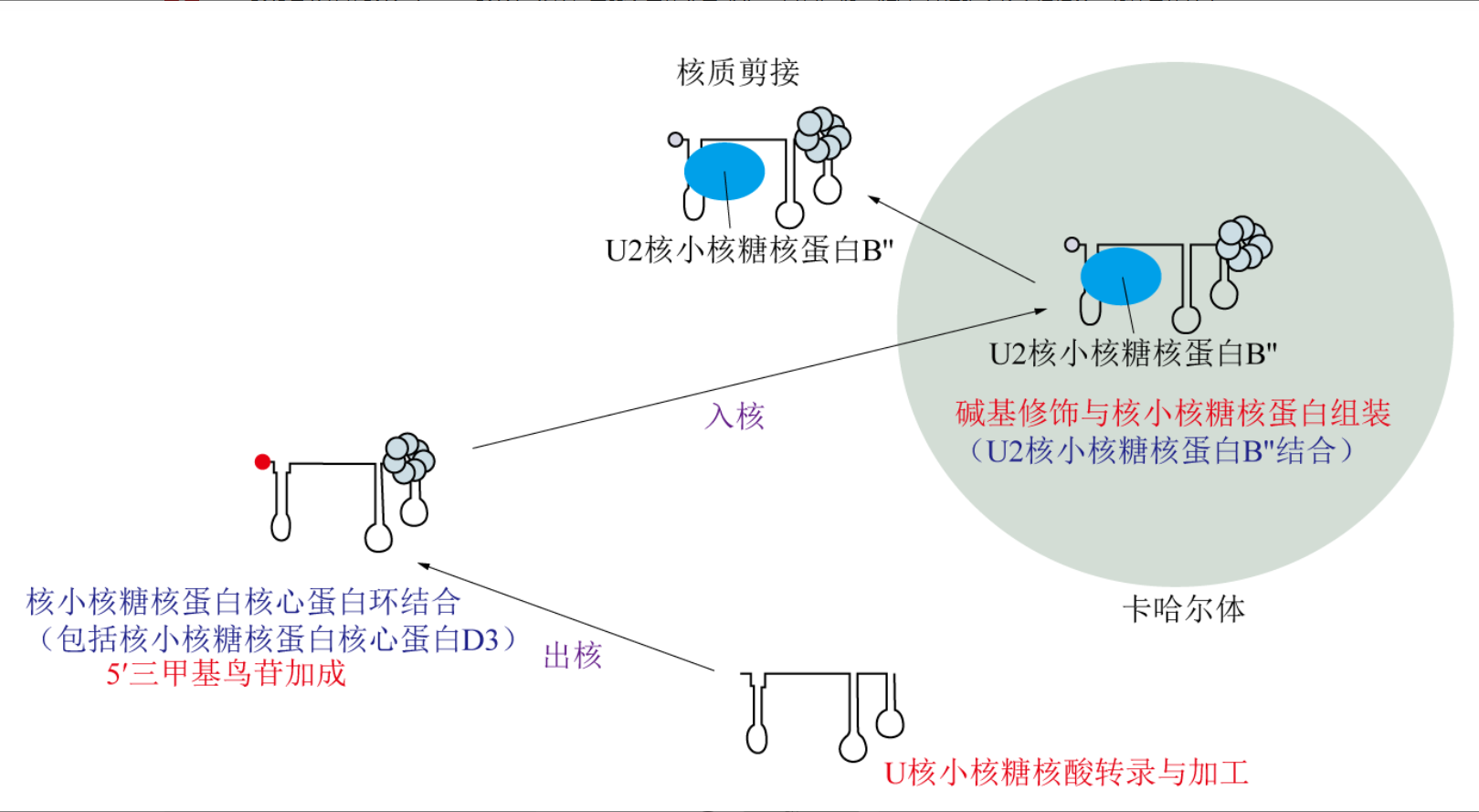 图1 卡哈尔体在snRNP成熟加工过程中的作用
图1 卡哈尔体在snRNP成熟加工过程中的作用

snoRNP组装与RNA修饰
核仁小核糖核蛋白是核糖体RNA加工所必需的复合物。纤维蛋白与NOPP140分别结合C/D box和H/ACA box RNA组分,在卡哈尔体中参与snoRNP的组装和功能调节。
ADSFAEQWER353423413434
组蛋白基因调控与端粒维持
卡哈尔体与染色体端粒的合成以及组蛋白基因簇的表观遗传修饰密切相关。端粒酶复合体在卡哈尔体中定位,维持端粒DNA合成所需酶的完整性。另外,通过调控组蛋白mRNA的3’端加工,卡哈尔体对染色质结构和表观遗传状态产生影响,进而调节细胞周期进程。 ADFASDFAF23RQ23R
p53信号通路交互
卡哈尔体还参与DNA损伤应激下的细胞周期检查点调控。紫外线照射可诱导其数量和分布形态改变。Coilin蛋白也被发现与p53信号通路的多种组分存在交互作用,在DNA损伤应答中协调细胞生长停滞与修复程序。 ADSFAEQWER353423413434
保守性与不同物种中的卡哈尔体
卡哈尔体在进化上是高度保守的核细胞器。到目前为止,在脊椎动物、果蝇、酵母菌以及植物中均发现存在卡哈尔体。果蝇的卡哈尔体已被鉴定并进行了研究,揭示了不同物种间RNA加工机制亚核组织的有趣差异。在裂殖酵母中,Coilin同源物Mug174同样能够通过相分离形成Cajal body,并且参与调控细胞在营养物质匮乏下的静息期维持。在植物中,卡哈尔体还具有物种特有的功能,包括通过NRPD1a RNA加工形成siRNA前体,以及RDR2/DCL3酶促反应生成表观调控复合物。 ADSFAEQWER353423413434
与疾病的关联
脊髓性肌萎缩症
脊髓性肌萎缩症(SMA)是一种常染色体隐性遗传的神经退行性疾病,由SMN1基因的纯合缺失或突变导致SMN蛋白水平降低而引起。SMN蛋白是Gemins复合体的关键组分,在snRNP组装中发挥核心作用。SMN缺失会导致神经元卡哈尔体的破坏。研究表明,I型SMA运动神经元中,每个神经元中卡哈尔体的平均数量严重减少,且大多数卡哈尔体无法招募SMN和剪接体snRNP,导致snRNP生物发生障碍和pre-mRNA剪接缺陷。SMN缺失还会导致Coilin蛋白异常地定位于多个核内病灶和核仁中,而非标准的卡哈尔体中。 ADFASDFAF23RQ23R
肿瘤发生
卡哈尔体的功能和表现异常也与肿瘤的发生相关。由于卡哈尔体与细胞周期、细胞增殖和应激反应等生物学过程密切相关,其在癌细胞中的形态和数量变化可能成为肿瘤诊断的潜在指标。
ADFASDFAF23RQ23R
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。