发育系统漂移
概念与提出
发育系统漂移(Developmental System Drift, DSD)由John R. True和Eric S. Haag于2001年正式命名,用于描述同源性性状的背后发育机制随进化时间发生分化的现象。它的提出是对传统同源性观念的反思:在经典定义中,“同源性”仅指性状本身来源于共同祖先,并不预设发育机制也必然保守。然而,研究者常不自觉地假设保守表型意味着保守机制,DSD直接挑战了这一“直觉”。
ADSFAEQWER353423413434
“漂移”一词的选用意在强调:发育机制分化的具体细节往往由中性过程(如中性突变和遗传漂变)主导,而非直接由自然选择驱动。但部分DSD也可能受间接选择影响,例如通过优化系统的稳健性。 ADFASDFAF23RQ23R
 图1 DSD的概念原理示意图
图1 DSD的概念原理示意图
发生机制
DSD的发生需要一个关键前提:发育系统的稳健性。稳健性是指系统在基因突变或环境扰动下仍能产生正常发育结果的能力。当稳健性足够高时,系统内存在缓冲和代偿机制,使得某些组分可以在不改变最终表型的前提下发生变化。
ADFASDFAF23RQ23R
顺-反式共进化是DSD最核心的机制之一:顺式调控元件和与其结合的转录因子(反式因子)在进化中发生补偿性相互变化——一方改变,另一方随之调整,从而维持系统输出不变。
ADSFAEQWER353423413434
线虫阴门发育的经典案例:秀丽隐杆线虫(C. elegans)和Pristionchus pacificus的阴门在最终形态上高度相似,但诱导信号通路不同(前者依赖EGF,后者依赖Wnt),部分信号分子调控逻辑甚至发生了反转。这展示了DSD在分子层面的具体表现。
ADFASDFAF23RQ23R
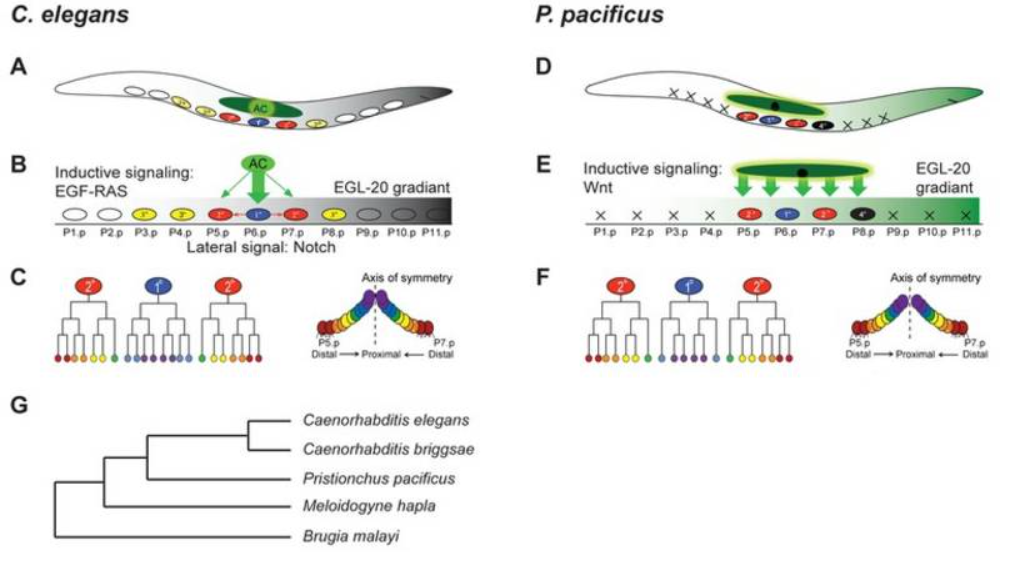 图2 太平洋拟盘龙线虫阴门发育案例示意图
图2 太平洋拟盘龙线虫阴门发育案例示意图
研究证据
- 线虫:对C. elegans和C. briggsae中超过1300个保守基因的RNAi比较发现,约25%的直系同源基因在不同物种中具有不同的体内功能,一些基因在一种物种中是必需的,在另一种中则非必需。
- 脊椎动物颅骨:蝾螈的颅骨发育模式与羊膜动物相似(可能是祖先状态),而非洲爪蟾的颅骨发育模式却发生了重大重排,但成体颅骨形态无明显差异。
- 海胆:形态几乎相同的海胆幼虫在不同物种中可由明显不同的调控网络构建而成。
- 昆虫定量系统漂移:果蝇与蚤蝇的间隙基因网络母源输入和动力学过程存在显著差异,但最终表达模式高度趋同。
进化意义
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。