发育扩展适应
定义与提出
发育扩展适应是演化生物学与发育生物学交叉融合的一个独特概念。“扩展适应”(Exaptation)一词由美国古生物学家Stephen Jay Gould和南非古生物学家Elisabeth Vrba于1982年正式提出。在此之前,这类现象一直被称为“预适应”(Preadaptation),但Gould和Vrba认为“预适应”一词容易让人误解为演化具有某种先见之明(仿佛自然演化可以“预见”未来),因此他们创造了“扩展适应”这一更中性的术语。 ADSFAEQWER353423413434
Gould和Vrba的定义可以简单概括为:一些最初具有其他用途、或者起初根本没有特定功能的生物特征,在演化的过程中被扩展到了当前的功能上。2001年,学者Chipman进一步提出了“发育扩展适应”的视角,将其聚焦于发育过程本身。他指出,发育程序的变化对生物体演化的影响,与外部环境的变化具有相似的重要作用。发育扩展适应是指在发育过程中预先存在的特征,在内外在环境的影响下产生了对某个特定环境生存有利的变化。
值得注意的是,Gould和Vrba在提出这一概念时,还进一步将扩展适应细分为两种情形:一种是从原有的适应结构中征用——即某个结构本来就是自然选择塑造出来的适应特征,后来又被用于其他功能;另一种是从非适应结构中征用——即某个结构在起源时并无特定适应功能,后来才偶然被利用起来。这两种情形的差异提醒我们,新功能的来源既可能是对现有适应结构的“二次利用”,也可能是对原本功能不明的“闲置部件”的偶然发掘。
ADSFAEQWER353423413434
 图1 发展扩展适应概念的演变过程
图1 发展扩展适应概念的演变过程
从达尔文的观察到现代理论
“特征在演化历史上可能发生功能上的变化”这一思想最早可以追溯到1859年达尔文在《物种起源》中的讨论。达尔文已经意识到,生物体的某个结构最初可能是为了解决某个问题而出现的,后来却被重新利用来解决完全不同的问题。 ADFASDFAF23RQ23R
在随后的一百多年里,这一现象被科学家称为“预适应”。这个词虽然沿用已久,但存在一个根本性缺陷:它暗示演化似乎有某种预见性——仿佛大自然能够提前进化出某个结构,“预见”到它将来会被用于别的目的。这种带有目的论色彩的表达对自然演化的科学描述造成了困扰。1982年,Gould和Vrba用“扩展适应”取代了“预适应”,并提出了一个更精确的“适应性扩展分类体系”,将生物特征按功能起源进行了更清晰的划分。
ADFASDFAF23RQ23R
2001年,Chipman将这一概念引入发育生物学领域,正式提出“发育扩展适应”。他关注的核心问题是:发育过程中那些在基因调控、细胞行为和组织分化等层面上预先存在的特征,如何在外界环境或内在发育压力的作用下,演化出有利于生物体生存的新功能。从发育角度来看,基因调控网络中的冗余连接、顺式调控元件的意外激活以及发育程序的偶然变异,都可能为新功能的出现提供“材料”。 ADSFAEQWER353423413434
典型案例
羽毛:从保温到飞行
羽毛是扩展适应最经典的范例。现代鸟类的羽毛高度复杂,具有羽轴、羽枝和小羽枝等精密结构,是飞行的关键装置。然而,大量化石证据表明,羽毛的演化历史远比飞行要久远得多。最早的羽毛化石见于不会飞行的兽脚类恐龙,其形态十分原始,呈中空圆柱形、没有分叉,主要功能很可能是维持体温。随着演化的推进,羽毛逐渐变得复杂——出现了分叉结构、形成了羽轴,之后又被用于展示和伪装等社交功能,最后才被用于飞行。支撑羽毛演化的基因调控网络也因此展现出高度的模块化特征,不同羽毛区域的专门化由不同的发育程序驱动,这种模块化结构使得羽毛能够逐步演化、不断积累新的功能。 ADSFAEQWER353423413434
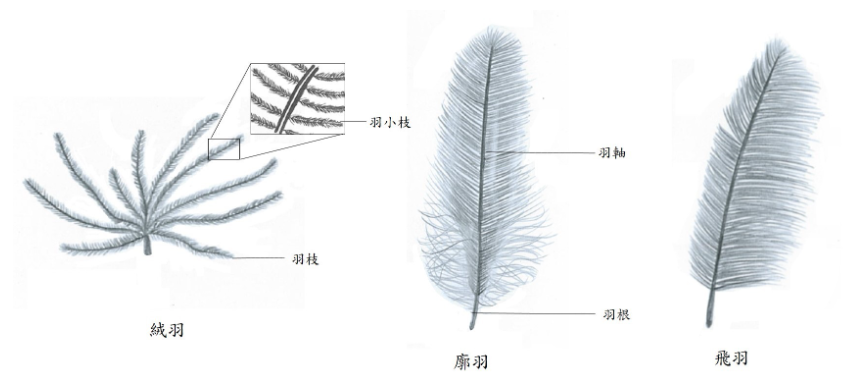 图2 羽毛演化扩展适应示意图
图2 羽毛演化扩展适应示意图
哺乳动物中耳:从咀嚼到听觉
哺乳动物中耳中的三块听小骨(镫骨、砧骨和锤骨)是人体骨骼系统中最小的骨头,形成了从鼓膜到内耳之间传递声波的听觉链。但它们的演化起源令人意想不到:锤骨和砧骨原本是爬行动物下颌中的关节骨和方骨,负责连接下颌与头骨、参与咀嚼运动。在哺乳动物演化过程中,这两块骨头逐渐与下颌分离、体积缩小并进入中耳,从“咀嚼参与者”变成了“听觉传递者”。这一转变是适应性扩展最生动的例证之一。值得注意的是,在这个过程中还存在一个过渡型中耳阶段——某些化石中这两块骨仍然通过软骨与下颌保持连接,反映了功能逐步转换的中间状态。
ADSFAEQWER353423413434
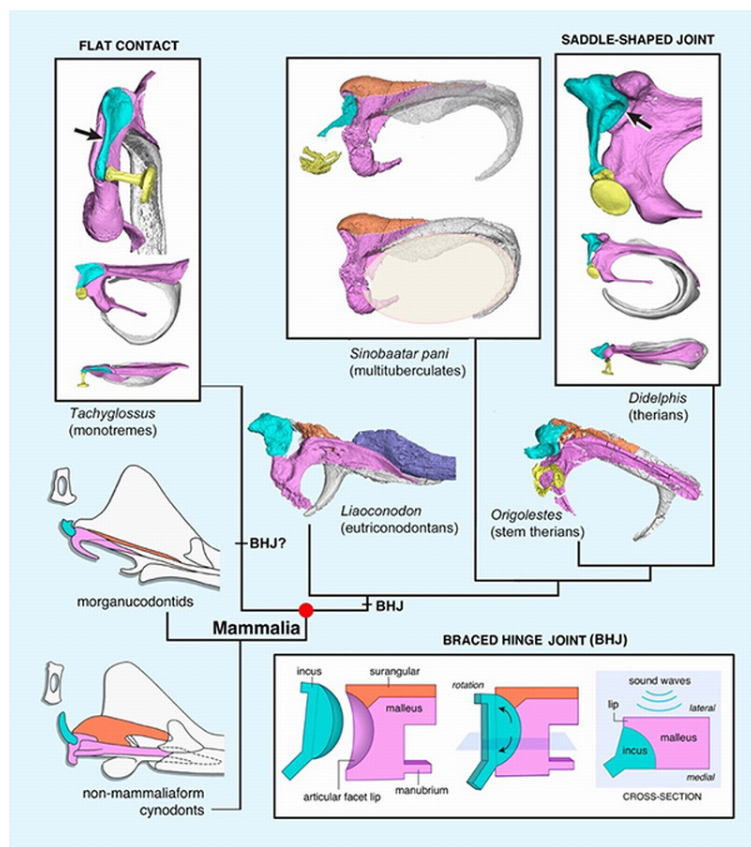 图3 哺乳动物中耳演化的祖先表型与过渡型固枢关节
图3 哺乳动物中耳演化的祖先表型与过渡型固枢关节
甲虫的角:从蜕皮工具到战斗武器
食粪金龟属(Onthophagus)甲虫的角提供了发育扩展适应的另一个生动例子。这些甲虫的角最初来源于幼虫期用于蜕皮的前胸角。在演化的过程中,这一原本只是辅助蜕皮的临时结构,逐渐被雄性甲虫用作争夺配偶的战斗武器,从而在性选择中占据了关键地位。这一转变体现了发育扩展适应的另一个重要特点:同一个发育程序可以被自然选择在不同的演化背景下反复“征用”,用于完全不同的生物学目标。
颈部肌肉:从鱼类到四足动物
2024年发表于《Nature Communications》的一项研究,从分子发育的角度揭示了颈部肌肉的起源。研究者通过比较胚胎发育中鱼类与四足动物(如两栖类、爬行类)的肌肉组来源发现,连接头部与躯干的肌肉群在两者之间具有保守的胚层起源。四足动物灵活的颈部并非从零开始构建,而是从鱼类祖先中已存在的肌肉组改造而来。这些肌肉被“征用”以获取适应陆地生活的新功能,支持了颈部扩展适应的发育基础。 ADFASDFAF23RQ23R
颅骨缝:从生长发育到分娩适应
所有脊椎动物幼崽的头骨上都有缝隙(颅骨缝),只有在哺乳动物中,这些缝隙被用来在分娩时允许头骨轻度变形,使婴儿更容易通过产道。原本适应于大脑快速生长而保留的临时性结构,被征用为解决分娩难题的“演化方案”。这种从“适应性特征”到“新的适应功能”的过渡,正是扩展适应理论所强调的核心模式。 ADSFAEQWER353423413434
翅鞘外骨骼:从飞行防护到抗旱铠甲
大多数甲虫用后翅飞行,而前翅硬化形成了保护性的鞘翅。但在铁甲虫(Phloeodes diabolicus)这个物种中,鞘翅已经永久锁定在一起,不再用于保护飞行翼,而是作为一种抗干燥的水分保持策略,帮助该物种适应干旱环境。这种将现有结构征用并加以改造的演化路径,进一步拓宽了我们对扩展适应多样性的认识。
分子层面的扩展适应
扩展适应并不仅限于宏观解剖结构,在分子层面同样普遍存在。基因组中大量的转座子(曾被称为“垃圾DNA”)就是最好的例子。这些可移动的DNA片段最初可能只是基因组中的寄生或随机片段,但随着演化的推进,它们被宿主基因组“驯化”,转变为具有重要功能的结构。例如,部分转座子贡献了转录因子结合位点和增强子等调控元件,参与了基因表达的精细调控。人类基因组中至少31%的冗余增强子对来源于独立转座子的扩展适应,提示转座子的驯化在基因组功能网络的演化中扮演了重要角色。免疫系统中负责抗体多样性的V(D)J重组机制也被认为起源于一个古老的RAG转座子。这说明,分子层面的扩展适应是推动基因调控网络复杂化和演化创新的核心驱动力之一,而以前被视为“功能沉默”的基因组重复序列也因此被重新认识。 ADFASDFAF23RQ23R
科学意义与研究争议
扩展适应的提出对传统演化理论做出了重要修正。在Gould和Vrba之前,演化生物学家习惯于将所有生物特征都称为“适应”,仿佛它们的功能就是它们被演化出来的原因。扩展适应理论提醒我们,一个特征当前的功能不等于它的演化起源——不能“因为现在有用,所以它就是为了这个用途而演化出来的”,这就是历史解读中常见的倒因为果。
但扩展适应概念也存在争议。一些学者质疑该概念“缺乏可操作性”——在具体的实证研究中很难严格区分“适应”和“扩展适应”,因为两者在演化过程中往往难以截然分开。例如,一个结构在被扩展适应后,后续的自然选择仍会对其进行塑造和优化,使其最终成为高度特化的适应结构。羽毛既曾被扩展适用于飞行,同时又受到了后续自然选择的持续修饰,所以“适应”和“扩展适应”更像是演化历程中的连续光谱,而非非此即彼的二分法。针对这一挑战,有研究者提出了检验“适应vs.扩展适应”的六条实证研究路径,为区分二者提供了更为系统的操作框架。此外,由于一些经典案例(如羽毛)难以进行严格的回溯性实验验证,也加重了实证检验的难度。不过,随着对基因调控和发育机制研究的深入,以及量化方法和计算机建模的应用,科学家对扩展适应的理解正在从描述性逐渐走向机制性。 ADSFAEQWER353423413434
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。