幕式物种形成
概念与定义
在主流的新达尔文主义框架下,物种形成通常被理解为一次漫长的“马拉松”:种群中积累的微小突变经自然选择筛选,经数万年、数十万年乃至更长时间才积累到足以产生生殖隔离的程度。 ADFASDFAF23RQ23R
幕式物种形成则提出了一种截然不同的“冲刺”模式:生物的变异是“非偶然”且“非渐进”的,甚至只需要一个步骤便能形成新物种。这一概念的基本内核包含以下几个要点。 ADSFAEQWER353423413434
(1)单步至多步的快速转变:在极端情况下,新物种可能在一个世代内直接出现。这种转变所涉及的遗传变化幅度极大,远超常态下的种内变异(如一个碱基的置换或小片段缺失)。典型的案例是植物中的多倍体化——一个二倍体植物通过染色体组整体加倍,在子一代即可形成与亲本在细胞学上完全不同的四倍体个体,此即“瞬时物种形成”(instantaneous speciation)。 ADFASDFAF23RQ23R
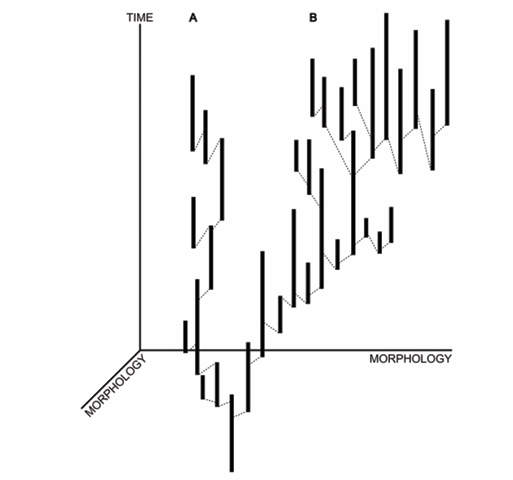
(2)跳越式的物种起源:幕式形成的新物种并非直接从祖先种“爬”出来,而是以“跳越”的方式出现在生态位或形态空间中的新位置。由于中间过渡形态极为罕见甚至可能不存在,这类物种在化石记录中会表现为突然出现,在间断平衡理论所讨论的“形态停滞期与快速变异的交替”中处于更加极端的维度。 ADFASDFAF23RQ23R
(3)与其他快速成种模式的边界区分: ADSFAEQWER353423413434
- 与地质尺度的快速事件的区别:生态系统在面临重大环境冲击(如小行星撞击、气候骤变)后,幸存的谱系会在数十万至数百万年内爆发式多样化(适应辐射)。但与幕式物种形成不同,适应辐射的驱动机制是生态位的空缺与自然选择,其“快速”是以地质时间为参照而言的,而非遗传意义上的瞬变。
- 与“有希望的怪物”假说的区别:1933年,遗传学家戈尔德施密特首次提出“有希望的怪物”一词,主张大型染色体结构重排(如染色体断裂与融合)可在单个世代中产生能够生殖隔离的极端变异体,从而启动新物种的诞生。戈尔德施密特本人并未提供令人信服的证据来解释这种“怪物”如何找到交配对象,使该假说长期备受冷落,但其核心思想与幕式物种形成高度重叠。
理论渊源与思想史
幕式物种形成是进化思想史上一场漫长论战的焦点之一。 ADFASDFAF23RQ23R
在查尔斯·达尔文之前,绝大多数学者相信物种可以经历突然而巨大的转变,这种观点被称为“骤变说”(Saltationism)。法国博物学家埃蒂安·若弗鲁瓦·圣伊莱尔是这一流派的激进代表,他在1820—1830年代提出,环境压力可诱导生物产生“怪物般”(monstrosities)的突变体,而这种突变体有可能成为新物种的“奠基者”。 ADSFAEQWER353423413434
1860年代,“跳跃式物种形成”进入了以弗朗西斯·高尔顿为代表的遗传学讨论。高尔顿在1869年的《遗传的天才》中首次表达了对此类事件的兴趣,并在1889年以后的著作中进一步将“骤变”视为一种可行的进化形式。 ADFASDFAF23RQ23R
达尔文之后,以德国胚胎学家威廉·希斯和动物学家威廉·贝特森为代表的“突变主义者”(Mutationists)坚持认为,物种形成中的关键步骤必然涉及一个或少数几个大效应突变,自然选择只能在突变出现之后对它们进行筛选,而非主动“塑造”它们。然而,20世纪30—40年代“现代综合进化论”(Modern Synthesis)的建立彻底改写了规则。该理论以遗传学为基石,将“微小突变+自然选择+隔离”作为物种形成的标准范式,而幕式物种形成因其难以用群体遗传学的数学框架解释,一度被边缘化为“非正统观念”。
遗传机制
尽管传统观点质疑其普遍性,但越来越多的现代研究表明,以下机制确实能够产生“准幕式”乃至“真正幕式”的物种形成事件。
3.1 多倍体化
多倍体化是幕式物种形成最确凿、可重复性最高的例证,常见于被子植物(如小麦、棉花、油菜、咖啡),在少数动物(如鲑科鱼类、一些两栖类)中也有零星报道。 ADFASDFAF23RQ23R
在自然界中,多倍体物种主要通过两种路径产生:一是同源多倍体(Autopolyploidy),即来自同一物种的不同个体之间发生基因组加倍;二是异源多倍体(Allopolyploidy),即不同物种之间杂交后再经染色体组加倍。后一过程尤为关键——杂交产生的杂合子在减数分裂中染色体无法正确配对,往往高度不育;但只要发生一次意外的染色体组加倍,全套染色体获得“备份”,不育性即可被完全解除,一个与两个亲本在遗传和生殖上均发生隔离的新物种几乎瞬间诞生。 ADSFAEQWER353423413434
多倍体化尤其擅长在短时间内组装新的性状组合:杂种F1代在基因组加倍后,双亲的整个基因套餐被一次性“封装”,由此产生的转录组大范围重塑可导致显著的表型分化,包括植株大小、抗逆性与次生代谢产物的质和量发生剧变。 ADFASDFAF23RQ23R
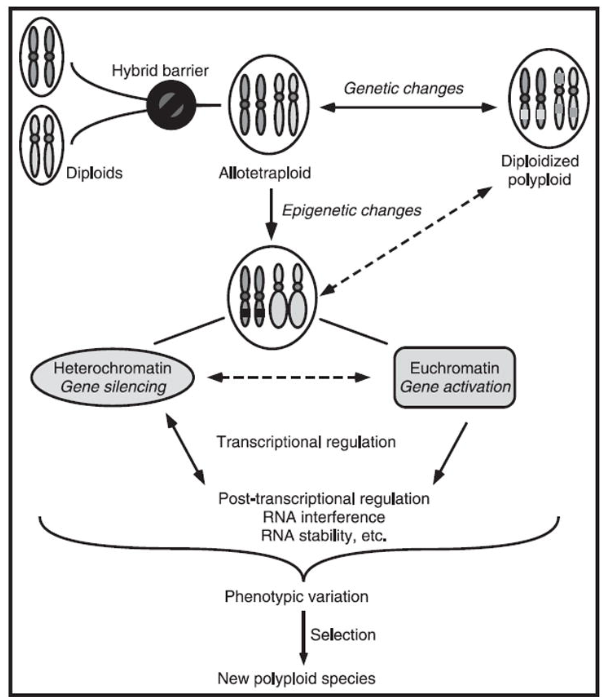 图2 异源多倍体的形成与演化示意图
图2 异源多倍体的形成与演化示意图
3.2 染色体裂变与融合
染色体的数目与结构并非一成不变。染色体裂变(一条整倍性染色体分裂为两条)和融合(两条合并为一条)可通过改变减数分裂同源配对的过程,直接引发生殖隔离。在物种形成率极高的蝴蝶类群(如绢蝶属Erebia)中,研究人员发现染色体数目的剧烈变化与物种多样性之间存在显著的正相关。中国科学家近年来在鼢鼠中鉴定出首个推动染色体裂变的分子机制——Aplf/Dna2基因的变异直接导致了一次精确的祖先染色体断裂事件,可能加速了整个支系的物种分化。
ADSFAEQWER353423413434
值得指出的是,在幕式物种形成的所有遗传机制中,染色体裂变与融合提供了一座微妙的桥梁:它不像多倍体化那样在一个世代内直接创造出全新物种,但可以显著加快种群分化,从而在“真正的大跳变”与“快速的小步积累”之间形成连续谱。
ADSFAEQWER353423413434
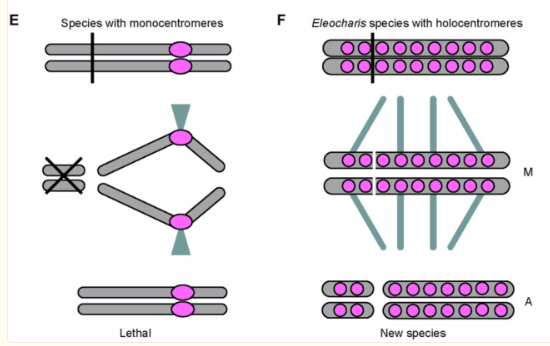
3.3 同倍体杂交物种形成
同倍体杂交物种形成是指杂交后代在染色体数目没有加倍(即仍保持二倍体水平)的条件下,通过重组和染色体结构重排固定一套独特的新基因型,使其与两个亲本均产生生殖隔离。向日葵中的沙漠向日葵(Helianthus paradoxus)即为一例——该物种的基因组一半来自普通向日葵,一半来自草原向日葵,但基因组合并后排除了有害重组产物,保留了特定的有利等位基因组合,使其在极端盐碱地带得以定殖并演化出新种。
ADFASDFAF23RQ23R
这一过程在“渐进”与“骤变”之间处于中间地带:它的产生依赖数次交叉与重组,故非多倍体那样的真正“瞬时事件”,但生殖隔离的形成仍可能远快于典型的异域物种形成。
3.4 系统内突变与“有希望的怪物”假说的再评价
戈尔德施密特曾推断,一次“系统突变”——即影响早期发育调控网络的大规模染色体重排——可能从单一个体中直接产生与亲代无法交配的罕见遗传类型,这些被自然选择判为“怪物”的个体,恰好有望成为新谱系的奠基者。在20世纪的大部分时间里,这一观点被主流学界嗤之以鼻,理由是“怪物”难以找到同类配偶。 ADSFAEQWER353423413434
现代群体遗传学为这一观点赋予了新的生命力:基因组学的进展显示,这种极端变异并非必须成对出现。如果一个携带着大效应突变的个体通过克隆繁殖(某些植物)、雌核生殖或与亲本有限度的回交产生的F2代在群体中稳定下来,那么“希望怪物”的类群就有可能成为现实。 ADSFAEQWER353423413434
化石记录中的快速成种事件
地质历史中蕴藏着大规模生物快速分化的丰富例证,可以归入“快速成种”而非“瞬时成种”的范畴,但在生物界对“幕式”一词的理解中,它们常常被视为重要的引述。
ADFASDFAF23RQ23R
在古生代奥陶纪大辐射事件中,腕足动物门在数百万年内出现了快速的分支演化,与显著的全球气候变化(海洋温度下降、大气氧含量升高、海平面剧烈振荡)精确对应。在更晚近的中新世—更新世,东非的丽鱼科鱼类上演了一场惊人的“适应辐射”——维多利亚湖的数百个丽鱼物种在不过十余万年的时间里快速演化出来,被认为是脊椎动物中已知最快的大规模物种辐射事件,其驱动因素包括生态位空缺、性选择以及可能发生的染色体易位和基因渗入性杂交。 ADFASDFAF23RQ23R
在这些辐射事件中,幕式物种形成可能并非唯一的引擎,而是在一定条件下与渐进演化共同推动生物多样性的形成。 ADSFAEQWER353423413434
学术争议与概念辨析
自20世纪70年代以来,古生物学家尼尔斯·埃尔德雷奇与史蒂芬·杰伊·古尔德提出的“间断平衡理论”持续引发关于进化节奏的激烈争论。该理论认为物种在绝大对数时间内处于表型停滞期(stasis),而成种事件集中出现在相对短暂的地质窗口内。
然而,一个长期存在的误解是将间断平衡与戈尔德施密特式的“有希望的怪物”直接等同。事实上,古尔德本人及其合作者多次强调,间断平衡所涉及的“快速”成种仍是在数万年的尺度上通过新达尔文主义机制(群体遗传漂变与自然选择)实现的,绝不等同于真正的“瞬变成种”。二者的混淆令“骤变”一词充满了争议色彩,使主流进化生物学家对一切暗示“跳跃式演化”的观点保持高度警惕。即便经历如此,许多学者仍承认,多倍体和染色体断裂所驱动的物种形成已构成了“真正幕式”的情形,至少在植物界和部分动物类群中不可否认。 ADSFAEQWER353423413434
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。