基因组编辑
基因组编辑技术总览编辑本段
基因组编辑
基因组编辑(Genome editing)泛指一切通过在特定基因组位点引入定向突变或修饰,从而改变生物体遗传信息的技术体系。其基本原理是利用工程化核酸酶在目标DNA序列处产生双链断裂(DSB),进而激活细胞内部的DNA损伤修复机制,实现遗传信息的精确改写。按照修复途径的不同,可分为依赖非同源末端连接(NHEJ)的随机插入或缺失(indel)突变,以及依赖同源定向修复(HDR)的精确模板插入或替换。早期技术如锌指核酸酶(ZFNs,1996年)、转录激活因子样效应物核酸酶(TALENs,2010年)通过人工设计蛋白结构域识别DNA序列,开创了靶向基因修饰的先河。2012年,CRISPR-Cas9系统的问世标志着基因组编辑迈入简易、高效、可编程的新时代。该系统利用单个引导RNA(sgRNA)与Cas9核酸酶形成复合体,通过sgRNA的20nt序列与靶DNA的Watson-Crick碱基配对实现识别,并识别原间隔序列邻近基序(PAM)以激活切割。此后,CRISPR-Cas12a(Cpf1)、Cas13(靶向RNA)等多样化酶系统的发现,以及dCas9融合效应蛋白(如转录调节因子、碱基修饰酶)的开发,极大丰富了编辑工具箱。2016年David Liu团队报道的碱基编辑器(Base Editors)利用失活Cas9融合脱氨酶,在无需DSB和供体模板条件下直接实现C·G→T·A或A·T→G·C的碱基转换,显著降低了调控风险。2019年开发的先导编辑器(Prime Editors)则通过Cas9切口酶融合逆转录酶,结合prime editing guide RNA(pegRNA)直接刻入新序列,实现了小片段插入、删除和任意碱基置换。当前,基因组编辑已覆盖从单细胞到模式生物、从体外细胞系到体内组织的多维度应用。 ADFASDFAF23RQ23R
核心分子机制与DNA修复途径编辑本段
基因组编辑的效能高度依赖内源性DNA修复通路的选择。NHEJ是细胞修复DSB的主要机制,尤其在G1期细胞中占主导地位。该途径将断裂末端直接连接,常导致1-50bp的插入或缺失(indel),从而造成基因移码或提前终止,适用于基因敲除。HDR则发生于S/G2期,利用同源模板(如外源供体DNA)进行精准修复,可实现特定点突变、基因标签插入或大片段替换。为提高HDR效率,研究者开发了抑制NHEJ的关键因子(如DNA-PKcs抑制剂)或细胞周期同步化策略。此外,微同源介导的端接(MMEJ)和单链退火(SSA)等替代修复途径也可被利用。碱基编辑器则绕开DSB,通过催化脱氨反应实现碱基直接转化,其机制不依赖NHEJ或HDR,从而避免随机indel的引入。先导编辑器通过Cas9切口酶在靶链产生切口,并由逆转录酶以pegRNA为模板合成新链,随后细胞修复机制整合该序列,实现精准编辑。这些差异化机制提供了从随机敲除到单碱基替换的阶梯式精准度。 ADSFAEQWER353423413434
关键技术方法对比编辑本段
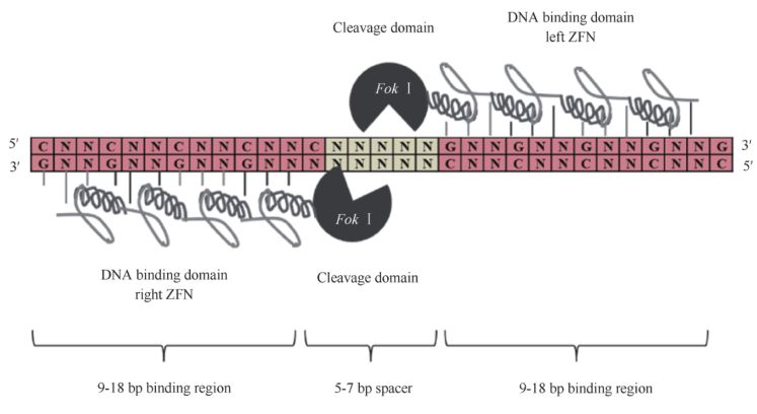
ZFNs由定制锌指蛋白串联体(每个识别3-4bp)与FokI核酸酶结构域组成,需二聚化产生DSB,设计耗时且脱靶效应显著。TALENs利用转录激活因子样效应物模块(每个识别单碱基)融合FokI,组装更为简便,但仍需蛋白质工程。CRISPR-Cas9以其RNA引导的简易性胜出,只需更改sgRNA序列即可重靶向,且可多重编辑。但Cas9的PAM序列(5'-NGG-3')限制靶位选择,脱靶效应尤其在富含GC区域明显。酶工程改进包括高保真Cas9变体(eSpCas9、SpCas9-HF1)和宽PAM变体(xCas9、SpRY)。CRISPR-Cas12a识别富含T的PAM(5'-TTN-3'),产生粘性末端,有利于NHEJ诱导大片段删除。碱基编辑器实现单碱基转换,但受制于窗口范围(~4-6nt)和旁侧序列偏好,可能产生脱靶编辑。先导编辑器虽理论上可任意替换,但效率较低(通常<30%),且对靶位环境敏感。在递送方面,质粒DNA、mRNA、核糖核蛋白(RNP)复合物及病毒载体(AAV、慢病毒、腺病毒)各有利弊:RNP方式瞬时且低脱靶,但成本高;AAV适合体内长期表达但包装容量小。
应用领域与里程碑编辑本段
在基础研究中,基因组编辑加速了基因功能鉴定、疾病模型构建及全基因组筛选。通过CRISPR文库筛选,已鉴定出若干肿瘤耐药基因(如WRN、POLE)和病毒宿主因子(如ACE2)。农业育种方面,基因编辑作物如抗锈病小麦、高油酸大豆已获批上市,并因不引入外源DNA而被部分国家豁免转基因监管。临床治疗领域,2023年底全球首款CRISPR基因编辑疗法Casgevy(exagamglogene autotemcel)获批用于镰刀型细胞贫血病和β地中海贫血,通过敲除BCL11A基因增强胎儿血红蛋白表达。此外,针对杜氏肌营养不良、视网膜色素变性、HIV潜伏库清除等数十项临床试验正在开展。合成生物学中,基因组编辑用于构建定制化微生物细胞工厂,如高产青蒿素的酵母菌株。在诊断方面,基于CRISPR效应酶(Cas12、Cas13)的特异性核酸快速检测平台(SHERLOCK、DETECTR)已用于新冠病毒、HPV及寨卡病毒检测。
技术局限与挑战编辑本段
当前基因组编辑面临多重瓶颈。脱靶效应可导致基因组不稳定和致癌风险,尤其在体内治疗中需严格评估。Cas9的PAM限制仍限制可靶向位点范围。递送困难是体内应用的主要障碍:AAV载体易引起免疫反应且靶向性有限,脂质纳米颗粒(LNP)虽有效但肝外递送效率低。碱基编辑器和先导编辑器虽提升了精准度,但仍有旁观者编辑(bystander editing)和pegRNA设计复杂等问题。此外,HDR在非分裂细胞中效率极低,限制了体细胞基因修复。伦理与监管问题同样突出,包括人类胚胎编辑的潜在滥用以及基因驱动对生态系统的不可逆影响。中国科学家贺建奎事件更警示需建立国际共识框架。 ADSFAEQWER353423413434
未来发展方向编辑本段
下一代技术将聚焦于开发超精准、低脱靶、广PAM的酶系统,如通过噬菌体辅助进化或深度突变筛选新型Cas变体。碱基编辑与先导编辑的优化将提升效率和特异性,并扩展编辑范围(如C·G到G·C的转换)。表观遗传编辑工具(如dCas9融合甲基转移酶或组蛋白修饰酶)可实现可逆基因调控而无DNA序列改变。在递送方面,新型工程化病毒衣壳(如AAV经定向进化靶向特定组织)和非病毒载体(如病毒样颗粒VLPs、外泌体)将推动体内应用。基因编辑联合其他疗法(如CAR-T细胞编辑、抗病毒基因沉积)将拓展疾病治疗策略。最后,建立涵盖安全性、可逆性和伦理的全球治理体系是实现临床转化的前提。
ADSFAEQWER353423413434
参考资料编辑本段
- Jinek M, Chylinski K, Fonfara I, et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 2012, 337(6096): 816-821.
- Doudna JA, Charpentier E. The new frontier of genome engineering with CRISPR-Cas9. Science, 2014, 346(6213): 1258098.
- Komor AC, Kim YB, Packer MS, et al. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature, 2016, 533(7603): 420-424.
- Gaudelli NM, Komor AC, Rees HA, et al. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature, 2017, 551(7681): 464-471.
- Anzalone AV, Randolph PB, Davis JR, et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 2019, 576(7785): 149-157.
- Hsu PD, Lander ES, Zhang F. Development and applications of CRISPR-Cas9 for genome engineering. Cell, 2014, 157(6): 1262-1278.
- Pickar-Oliver A, Gersbach CA. The next generation of CRISPR–Cas technologies and applications. Nature Reviews Molecular Cell Biology, 2019, 20(8): 490-507.
- Ledford H. Landmark CRISPR therapy shows promise for sickle-cell disease. Nature, 2023, 624(7991): 22.
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。