碱基互补配对
碱基互补配对的化学本质与氢键作用编辑本段
 ADFASDFAF23RQ23R
ADFASDFAF23RQ23R 碱基互补配对
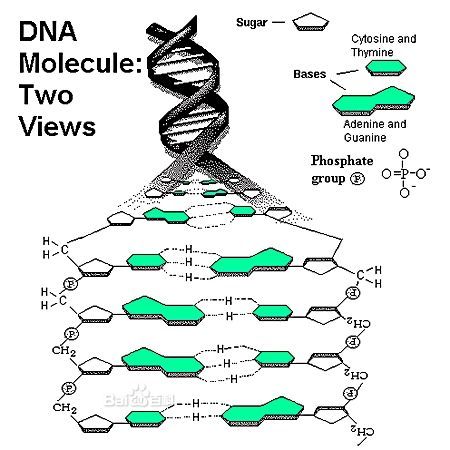
碱基互补配对(Complementary Base Pairing)的核心在于嘌呤与嘧啶碱基间定向形成的氢键网络。氢键是一种弱相互作用,但大量氢键的协同效应赋予DNA双螺旋高度稳定性。腺嘌呤(A)的N1与胸腺嘧啶(T)的N3之间形成一个氢键,同时A的N6H2与T的O4形成第二个氢键;鸟嘌呤(G)的O6与胞嘧啶(C)的N4H2形成第一个氢键,G的N1H与C的N3形成第二个,G的N2H2与C的O2形成第三个。这种一对一识别使得A-T/A-U(两个氢键)和G-C(三个氢键)成为唯二的标准配对模式。G-C配对的三个氢键赋予其更高的热稳定性,解链温度(Tm)显著高于A-T丰富的区域。
ADSFAEQWER353423413434
DNA双螺旋结构中的碱基配对几何约束编辑本段
在B型DNA双螺旋中,碱基平面垂直于螺旋轴,配对碱基位于螺旋内侧,形成堆积的疏水核心。碱基配对必须满足严格的几何参数:嘌呤与嘧啶的配对使得两条链间距离保持恒定(约2.0 nm),糖-磷酸骨架的扭曲形成大沟和小沟,分别用于蛋白质识别与特异性结合。大沟富含氢键供体与受体,是转录因子结合的关键位点;小沟则更窄,常用于小分子药物如纺锤菌素和偏端霉素的识别。Watson-Crick配对确保了双螺旋的互补链反向平行(5'→3'与3'→5'),是DNA半保留复制的结构前提。 ADSFAEQWER353423413434
RNA与DNA配对的差异及功能意义编辑本段
RNA中尿嘧啶(U)替代胸腺嘧啶(T),A-U配对仅形成两个氢键,相比A-T(也为两个氢键)热稳定性类似,但U的甲基缺失影响RNA双螺旋的构象柔性。RNA往往以单链形式存在,但在转运RNA(tRNA)和核糖体RNA(rRNA)中通过局部碱基配对形成三叶草结构和假结等二级结构。RNA-RNA配对在剪接体、端粒酶和CRISPR-Cas系统中发挥机械和催化功能;DNA-RNA杂交双链(R-loop)在转录和基因组稳定性调控中至关重要,其稳定性介于DNA双链和RNA双链之间。
ADFASDFAF23RQ23R
非标准碱基配对:摆动配对与化学修饰的影响编辑本段
标准Watson-Crick配对并非绝对,生物体内存在多种非标准配对。最著名的是Crick提出的摆动假说(Wobble Hypothesis):密码子第三位碱基与反密码子第一位碱基间的配对允许非严格识别,例如G与U、I(次黄嘌呤)与A、U、C。这种灵活性提高了遗传密码的容错性并减少了tRNA种类。此外,碱基化学修饰(如甲基化、羟甲基化)可改变配对特性:5-甲基胞嘧啶保留与G配对能力,但5-羟甲基胞嘧啶可能促进与T配对;N6-甲基腺嘌呤干扰与T配对。这些修饰在表观遗传调控和基因表达中起关键作用。 ADFASDFAF23RQ23R
热力学参数与DNA熔解动力学编辑本段
碱基配对的热力学由氢键(焓变ΔH)和堆积作用(熵变ΔS)共同决定。最近邻模型(Nearest-Neighbor Model)精确计算每个双链碱基对的ΔG°:A-T配对约1.0-1.3 kcal/mol,G-C配对约2.0-2.3 kcal/mol。连续G-C区域使DNA更耐高温,PCR引物的Tm值可用公式Tm = 4(G+C) + 2(A+T)估算(短链)或更为精确的Wallace公式。DNA熔点随GC含量线性增加,每增加1% GC约使Tm升高0.4°C。复性动力学研究表明,碱基配对的精确性通过分子碰撞和错配识别实现高保真度,错配引起的自由能增加(约1-4 kcal/mol)是DNA聚合酶校正活性的基础。 ADSFAEQWER353423413434
碱基配对在遗传信息传递中的核心作用编辑本段
DNA复制时,DNA聚合酶根据模板链选择互补的脱氧核苷三磷酸(dNTP),确保子链与母链反向互补。RNA转录时,RNA聚合酶以DNA为模板合成互补RNA,其中A对应U。翻译过程中,mRNA密码子与tRNA反密码子通过碱基配对(遵摆动假说)将核苷酸序列解码为氨基酸序列。碱基配对的保真性通过聚合酶的校对功能(3'→5'外切酶活性)和错配修复系统(如MutS-MutL-MutH)进一步强化,使突变率低于10⁻⁹每碱基每复制。逆转录病毒(如HIV)的逆转录酶也依赖碱基配对合成DNA前病毒。
生物技术中对碱基配对的利用与改造编辑本段
聚合酶链反应(PCR)利用引物与模板的严格配对扩增DNA;DNA微阵列依赖靶分子与探针的互补杂交检测基因表达;荧光原位杂交(FISH)通过探针定位染色体;CRISPR-Cas系统中,向导RNA(gRNA)与靶DNA通过Watson-Crick配对引导Cas9切割。在合成生物学中,人工碱基对(例如d5SICS-dNaM或P-Z)已被创建并引入生物体内,实现了六字母遗传系统,显著扩展了遗传密码。此外,锁核酸(LNA)和肽核酸(PNA)修饰可提高配对的亲和力和特异性,用于反义治疗和诊断探针设计。
ADSFAEQWER353423413434
碱基配对的进化起源与展望编辑本段
RNA世界假说认为,早期生命基于RNA的碱基配对实现自我复制。现代核酸世界保留了此核心机制,并演化出多种调控层次。未来,基于碱基配对的纳米技术,如DNA折纸和分子机器,将利用序列编码自组装三维结构;同时,碱基配对在基因治疗中引导反义寡核苷酸和siRNA靶向特定mRNA。理解碱基互补配对的细微之处是分子生物学和精准医学的根本基石。
ADSFAEQWER353423413434
参考资料编辑本段
- Watson JD, Crick FHC. Molecular structure of nucleic acids: a structure for deoxyribose nucleic acid. Nature. 1953;171(4356):737-738.
- Crick FHC. Codon–anticodon pairing: the wobble hypothesis. J Mol Biol. 1966;19(2):548-555.
- SantaLucia J, Hicks D. The thermodynamics of DNA structural motifs. Annu Rev Biophys Biomol Struct. 2004;33:415-440.
- Kool ET. Hydrogen bonding, base stacking, and steric effects in DNA replication. Annu Rev Biophys Biomol Struct. 2001;30:1-22.
- Malyshev DA, Dhami K, Lavergne T, et al. A semi-synthetic organism with an expanded genetic alphabet. Nature. 2014;509(7500):385-388.
- Jinek M, Chylinski K, Fonfara I, et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 2012;337(6096):816-821.
- Saenger W. Principles of Nucleic Acid Structure. Springer-Verlag; 1984.
- Sinden RR. DNA Structure and Function. Academic Press; 1994.
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。