A-β蛋白
β-淀粉样蛋白(Amyloid-β,Aβ)
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
基础信息
ADSFAEQWER353423413434
ADSFAEQWER353423413434
| 项目 | 内容 | ADFASDFAF23RQ23R
| 中文名 | β-淀粉样蛋白 |
| 英文名 | Amyloid-β(Aβ) |
ADSFAEQWER353423413434
| 别名 | β-淀粉样肽、Aβ肽、Aβ蛋白 |
ADSFAEQWER353423413434
| 分子量 | 约 4.5 kDa | ADFASDFAF23RQ23R
| 前体蛋白 | 淀粉样前体蛋白(APP) |
| 产生途径 | APP 经 β-和 γ-分泌酶协同切割 |
ADFASDFAF23RQ23R
| 主要亚型 | Aβ40、Aβ42 | ADSFAEQWER353423413434
| 所属学科 | 神经生物学、分子生物学、病理学 | ADFASDFAF23RQ23R
| 相关疾病 | 阿尔茨海默病、唐氏综合征、脑淀粉样血管病 |
ADSFAEQWER353423413434
 ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
一、定义编辑本段
β-淀粉样蛋白(Amyloid-β,简称 Aβ)是由 36-43 个氨基酸组成的小分子多肽,分子量约为 4.5 kDa。它是淀粉样前体蛋白(Amyloid Precursor Protein,APP)经 β-分泌酶和 γ-分泌酶顺序切割后产生的蛋白水解片段。
ADSFAEQWER353423413434
ADSFAEQWER353423413434
Aβ 是阿尔茨海默病患者大脑中淀粉样斑块的主要成分,其异常聚集被认为是阿尔茨海默病发病的核心事件之一。在正常生理条件下,Aβ 的产生与清除处于动态平衡;当这一平衡被打破时,Aβ 发生错误折叠和异常聚集,形成从可溶性寡聚体到不溶性淀粉样纤维的多种聚集体,最终沉积为老年斑。 ADFASDFAF23RQ23R
ADSFAEQWER353423413434
ADSFAEQWER353423413434
二、发现历史编辑本段
ADSFAEQWER353423413434
(一)淀粉样斑块的早期描述
1906 年,德国神经病理学家阿洛伊斯·阿尔茨海默(Alois Alzheimer)在解剖一名早发性痴呆患者的大脑时,首次描述了两种特征性的病理结构:**淀粉样斑块**(老年斑)和**神经原纤维缠结**。然而,当时并不清楚这些斑块的化学组成。
(二)Aβ 的鉴定
ADFASDFAF23RQ23R
1984 年,科学家首次从阿尔茨海默病患者大脑的淀粉样斑块中分离并鉴定了 Aβ 肽的氨基酸序列,确认其为一种由约 40 个氨基酸组成的多肽。这一发现开启了以 Aβ 为核心的阿尔茨海默病研究新纪元。 ADFASDFAF23RQ23R
ADSFAEQWER353423413434
(三)APP 基因的克隆与 Aβ 起源的阐明
1987 年,APP 基因被成功克隆。随后的研究表明,Aβ 并非一个独立的基因产物,而是由 APP 经蛋白水解切割产生。这一发现将 Aβ 的产生从“神秘来源”拉回到了明确的分子途径上,为后续的药物开发奠定了基础。 ADFASDFAF23RQ23R
ADSFAEQWER353423413434
(四)淀粉样蛋白级联假说的提出
1992 年,John Hardy 和 Gerald Higgins 正式提出了 **“淀粉样蛋白级联假说”** (Amyloid Cascade Hypothesis)。该假说认为,Aβ 的异常聚集是触发阿尔茨海默病一系列病理事件(包括 tau 蛋白过度磷酸化、神经炎症、突触功能障碍和神经元死亡)的始动因素。这一假说在此后三十年间主导了 AD 的研究和药物开发方向。
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
三、Aβ 的产生编辑本段
ADSFAEQWER353423413434
(一)前体蛋白:淀粉样前体蛋白(APP)
ADFASDFAF23RQ23R
APP 是一种 I 型跨膜糖蛋白,分子量约 100–140 kDa,在全身多种组织中均有表达,尤其在神经元的突触部位表达丰富。APP 参与神经元发育、信号转导、细胞内运输等多种生物学功能。2026 年的一项研究还发现,APP 可能通过溶酶体胞吐主动保护神经元免受核损伤。
ADSFAEQWER353423413434
APP 的代谢命运取决于首先切割它的酶——它面临两条截然不同的加工途径。 ADFASDFAF23RQ23R
ADSFAEQWER353423413434
(二)非淀粉样蛋白生成途径(健康途径)
ADSFAEQWER353423413434
在健康个体中,APP 主要通过 **α-分泌酶** 在 Aβ 序列内部进行切割。由于切割位点位于 Aβ 结构域的中段,这一过程**直接破坏了完整 Aβ 肽段的形成**,同时产生具有神经营养活性的可溶性 APPα(sAPPα)片段。此途径是健康细胞中 APP 加工的主要方式。
(三)淀粉样蛋白生成途径(致病途径)
在病理条件下,APP 先后被 **β-分泌酶**(BACE1)和 **γ-分泌酶** 切割: ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
1. β-分泌酶切割:在 APP 的胞外区域靠近跨膜域的位置进行切割,产生一个可溶性的 N 端片段(sAPPβ)和一个膜结合的 C99 片段。
ADSFAEQWER353423413434
2. γ-分泌酶切割:在跨膜域内部对 C99 进行切割,释放出 Aβ 肽段。
ADFASDFAF23RQ23R
γ-分泌酶的切割位点并不完全固定,因此会产生不同长度的 Aβ 肽段。
ADSFAEQWER353423413434
ADFASDFAF23RQ23R
(四)Aβ 的主要亚型
ADSFAEQWER353423413434
| 亚型 | 氨基酸长度 | 特点 | ADSFAEQWER353423413434
| Aβ40 | 40 个氨基酸 | 含量最丰富(约占 80-90%),溶解度较高 | ADSFAEQWER353423413434
| Aβ42 | 42 个氨基酸 | 含量较少但疏水性更强,更易聚集 | ADFASDFAF23RQ23R
| Aβ43 | 43 个氨基酸 | 含量极少 | ADFASDFAF23RQ23R
Aβ42 因具有更强的疏水性和更快的聚集速度,被认为是更具神经毒性的亚型,在阿尔茨海默病的发病中扮演更为关键的角色。
ADSFAEQWER353423413434
四、Aβ 的清除编辑本段
Aβ 的清除与产生同等重要。Aβ 在大脑中的浓度由**产生速率**和**清除速率**共同决定。 ADSFAEQWER353423413434
Aβ 的清除主要通过以下途径实现: ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
| 清除途径 | 机制 |
| 酶降解 | 多种蛋白酶(如胰岛素降解酶 IDE、脑啡肽酶、ACE 等)可降解 Aβ |
ADSFAEQWER353423413434
| 血脑屏障转运 | 通过 LRP-1 等转运体将 Aβ 从脑内泵出至外周循环 |
ADFASDFAF23RQ23R
| 小胶质细胞吞噬 | 小胶质细胞可吞噬并清除 Aβ 沉积物 |
| 脑脊液循环 | Aβ 可通过脑脊液循环被带走 | ADFASDFAF23RQ23R
ADSFAEQWER353423413434
当 Aβ 的产生超过清除能力时(如衰老过程中清除效率下降),Aβ 便会在脑内逐渐累积。
ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
五、Aβ 的聚集与结构编辑本段
ADFASDFAF23RQ23R
(一)聚集过程的阶段性
ADSFAEQWER353423413434
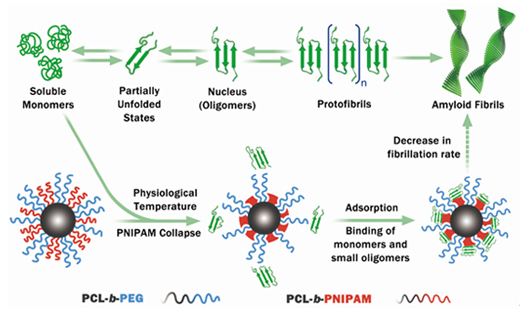
Aβ 单体在特定条件下会经历多步聚集过程,形成结构和毒性各异的聚集体: ADFASDFAF23RQ23R
ADSFAEQWER353423413434
```
ADSFAEQWER353423413434
Aβ 单体 → 可溶性寡聚体 → 原纤维 → 成熟纤维 → 淀粉样斑块
```
ADFASDFAF23RQ23R
Aβ 的错误折叠和聚集是一个高度复杂的过程,涉及多种中间体。
ADFASDFAF23RQ23R
(二)各聚集阶段的特征
ADSFAEQWER353423413434
| 聚集形式 | 结构特征 | 毒性特征 |
ADFASDFAF23RQ23R
| Aβ 单体 | 无规卷曲或 α-螺旋构象 | 无明显毒性 |
ADFASDFAF23RQ23R
| 可溶性寡聚体 | 由数个至数十个单体组成 | **毒性最强**的 Aβ 形式,是强效突触毒素 |
ADFASDFAF23RQ23R
| 原纤维 | 中间状态,β-折叠含量增加 | 具有一定毒性 | ADSFAEQWER353423413434
| 成熟纤维 | 高度有序的交叉 β-折叠结构 | 构成斑块的主要成分,毒性相对较低 |
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
(三)结构特征
ADSFAEQWER353423413434
Aβ 纤维的核心结构为 **交叉 β-折叠**(cross-β-sheet),其中 β-链垂直于纤维长轴排列。Aβ42 纤维通常由每层两个分子组成,残基 15–42 参与形成核心结构。Aβ 具有高度的构象可塑性,可根据环境条件采用无规卷曲、螺旋或 β-发夹等多种结构。
ADFASDFAF23RQ23R
六、Aβ 的毒性机制编辑本段
ADFASDFAF23RQ23R
(一)直接神经毒性
ADSFAEQWER353423413434
Aβ 寡聚体和斑块通过多种机制损伤神经元:
ADFASDFAF23RQ23R
- 突触毒性:Aβ 寡聚体是强效的**突触毒素**,可直接损害突触结构和功能
- 线粒体损伤:抑制线粒体活性,影响能量代谢 ADSFAEQWER353423413434
ADFASDFAF23RQ23R
- 炎症激活:刺激神经炎症过程 ADFASDFAF23RQ23R
ADSFAEQWER353423413434
(二)间接致病机制
ADFASDFAF23RQ23R
除了直接毒性,Aβ 还通过以下间接途径参与疾病进程:
ADSFAEQWER353423413434
ADFASDFAF23RQ23R
- tau 蛋白病理:Aβ 可改变激酶和磷酸酶活性,导致 tau 蛋白过度磷酸化并形成神经原纤维缠结 ADSFAEQWER353423413434
- 血脑屏障破坏:Aβ 可损害血脑屏障的完整性
- 自噬与线粒体功能障碍:干扰细胞的自噬清除系统和线粒体功能 ADSFAEQWER353423413434
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
七、Aβ 的生理功能编辑本段
长期以来 Aβ 被视为纯粹的“病理蛋白”,但近年研究表明 Aβ 在正常生理条件下可能具有重要的生物学功能。 ADSFAEQWER353423413434
ADFASDFAF23RQ23R
Aβ 的潜在生理功能包括:
ADFASDFAF23RQ23R
- 促进神经元生长
- 稳定和组装微管
- 促进轴突伸长
ADSFAEQWER353423413434
- 调节突触传递和可塑性 ADSFAEQWER353423413434
Aβ 正常生理功能的丧失也被认为是导致神经元功能障碍的原因之一。Aβ 的“生理 vs 病理”双重角色使其成为一个复杂的研究对象。
ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
ADSFAEQWER353423413434
八、Aβ 相关疾病编辑本段
ADFASDFAF23RQ23R
(一)阿尔茨海默病(核心关联)
ADSFAEQWER353423413434
阿尔茨海默病是最常见的痴呆类型,其特征性病理改变包括 **β-淀粉样蛋白沉积** 和 **神经原纤维缠结**。在患者大脑中,淀粉样前体蛋白的处理发生改变,导致 β-淀粉样纤维状沉积和聚集。
流行病学数据: ADFASDFAF23RQ23R
- 阿尔茨海默病占老年痴呆病例的 60%–80%
- 在美国,约 11% 的 ≥65 岁人群患有 AD
ADFASDFAF23RQ23R
- 65–74 岁:约 5%;75–84 岁:约 13%;≥85 岁:约 33%
- 女性发病率约为男性的 2 倍
ADSFAEQWER353423413434
ADFASDFAF23RQ23R
(二)其他相关疾病
ADFASDFAF23RQ23R
- 唐氏综合征:由于 21 号染色体三体(APP 基因位于 21 号染色体),患者早年即可出现 Aβ 沉积和 AD 样病理
ADSFAEQWER353423413434
- 脑淀粉样血管病(CAA) :Aβ 沉积于脑部血管壁,可导致血管功能障碍和出血风险 ADFASDFAF23RQ23R
- 包涵体肌炎:Aβ 在肌肉组织中的异常沉积 ADSFAEQWER353423413434
ADSFAEQWER353423413434
ADFASDFAF23RQ23R
九、Aβ 的检测方法编辑本段
(一)影像学检测
| 检测方法 | 原理 | 应用 | ADFASDFAF23RQ23R
| Aβ-PET | 使用 Aβ 特异性示踪剂(如 Pittsburgh Compound B)进行正电子发射断层扫描 | 活体显示脑内 Aβ 斑块分布 | ADSFAEQWER353423413434
| FDG-PET | 检测脑部葡萄糖代谢 | 评估神经元功能活动 |
ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
(二)体液生物标志物
- 脑脊液(CSF)检测:测量 Aβ42 水平及 Aβ42/Aβ40 比值。**Aβ42/Aβ40 比值下降**通常预示着大脑中淀粉样斑块的沉积 ADFASDFAF23RQ23R
- 血液检测:近年来基于血液的 Aβ 检测技术快速发展,可用于大规模筛查
(三)免疫学检测
ADFASDFAF23RQ23R
ELISA(酶联免疫吸附测定) 是定量检测 Aβ 水平的常用方法。通过抗原-抗体特异性结合,可检测脑组织匀浆、脑脊液、血浆等样本中的 Aβ 含量。近年来还开发了针对 Aβ 寡聚体的特异性免疫检测方法。
ADFASDFAF23RQ23R
ADFASDFAF23RQ23R
十、Aβ 靶向治疗策略编辑本段
ADSFAEQWER353423413434
(一)抗 Aβ 单克隆抗体(已获批)
ADFASDFAF23RQ23R
近年来,靶向 Aβ 的单克隆抗体疗法取得了突破性进展。 ADSFAEQWER353423413434
ADFASDFAF23RQ23R
| 药物名称 | 类型 | 作用机制 | 状态 |
ADFASDFAF23RQ23R
| 仑卡奈单抗(Lecanemab) | 抗 Aβ 单抗 | 清除 Aβ 斑块,延缓早期 AD 认知衰退 | 已获批 | ADFASDFAF23RQ23R
| 多奈单抗(Donanemab) | 抗 Aβ 单抗 | 清除 Aβ 斑块 | 已获批 |
ADSFAEQWER353423413434
这些抗 Aβ 单抗被归类为 **疾病修饰治疗**(Disease-Modifying Therapy),研究一致显示它们可以有效减少 PET 上的淀粉样斑块负荷。治疗适用人群主要为 **早期阿尔茨海默病患者**,需要严格的生物标志物分层。
ADSFAEQWER353423413434
ADFASDFAF23RQ23R
(二)其他 Aβ 靶向策略
ADSFAEQWER353423413434
| 策略 | 机制 | 说明 |
| β-分泌酶抑制剂 | 抑制 BACE1,减少 Aβ 产生 | 临床开发中,面临安全性挑战 | ADSFAEQWER353423413434
| γ-分泌酶抑制剂/调节剂 | 抑制或调节 γ-分泌酶活性 | 因影响 Notch 信号通路,面临毒性问题 |
| Aβ 聚集抑制剂 | 阻止 Aβ 单体聚集为有毒寡聚体 | 临床前/早期临床研究 | ADSFAEQWER353423413434
| Aβ 降解酶 | 增强 Aβ 的酶促降解 | 研究阶段 | ADSFAEQWER353423413434
| Aβ 疫苗(主动免疫) | 诱导机体产生抗 Aβ 抗体 | 部分候选药物进入临床 | ADSFAEQWER353423413434
| CAR-星形胶质细胞疗法 | 工程化细胞清除 Aβ | 前沿探索 |
ADFASDFAF23RQ23R
(三)治疗挑战与争议
ADSFAEQWER353423413434
尽管抗 Aβ 抗体在清除斑块方面效果显著,但其对 **临床结局的疗效相对有限**。部分研究指出,成功清除大脑中的 Aβ 蛋白与临床症状改善之间缺乏明确的相关性。这些争议促使研究者将神经炎症、线粒体功能障碍、氧化应激等机制纳入更全面的疾病模型。 ADSFAEQWER353423413434
ADSFAEQWER353423413434
ADSFAEQWER353423413434
十一、研究前景与挑战编辑本段
ADSFAEQWER353423413434
(一)主要科学挑战
ADFASDFAF23RQ23R
1. Aβ 假说的完善:Aβ 单抗临床疗效有限,促使学界重新审视和修正经典的淀粉样蛋白级联假说 ADSFAEQWER353423413434
2. 毒性形式的确定:究竟哪种 Aβ 形式(寡聚体、原纤维还是斑块)是主要的毒性实体,仍未完全明确
ADSFAEQWER353423413434
3. 生理功能的干扰:靶向 Aβ 的治疗可能干扰其正常生理功能 ADSFAEQWER353423413434
4. 治疗时间窗:何时开始干预最为有效(症状前 vs 症状期) ADSFAEQWER353423413434
ADSFAEQWER353423413434
(二)未来研究方向
ADSFAEQWER353423413434
- 早期诊断:开发更灵敏的血液 Aβ 生物标志物,实现症状前筛查 ADFASDFAF23RQ23R
- 联合治疗:将抗 Aβ 治疗与抗 tau、抗炎、代谢调节等策略相结合
ADSFAEQWER353423413434
- 新型降解技术:探索 PROTAC 等蛋白质降解技术在 Aβ 清除中的应用 ADSFAEQWER353423413434
ADFASDFAF23RQ23R
(三)Aβ 给予人类的启示
从 1906 年阿尔茨海默医生在显微镜下首次看到那些神秘的斑块,到 2020 年代抗 Aβ 抗体走入临床,β-淀粉样蛋白的研究历程跨越了整整一个世纪。Aβ 的故事是科学探索的缩影——它充满曲折和争议,既有理论的胜利(Aβ 假说主导 AD 研究三十年),也有现实的困境(临床疗效未达预期)。
Aβ 提醒我们,神经退行性疾病的本质可能是多因素、多通路交织的复杂网络,而非单一“坏蛋白”的简单故事。理解 Aβ 在生理与病理之间的微妙平衡,或许才是解开阿尔茨海默病之谜的关键钥匙。 ADSFAEQWER353423413434
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。