复杂细胞
定义与发现编辑本段
复杂细胞(complex cell)是视觉皮层中一类对方向性刺激具有选择性,但不受刺激位置精细变化影响的神经元。它们最早由David Hubel和Torsten Wiesel在1959年通过猫的视觉皮层电生理实验发现。这些细胞主要位于初级视皮层(V1)以及次级视皮层(V2、V3等),尤其在Brodmann 18区和19区高度集中。Hubel和Wiesel通过记录单个神经元的电活动,揭示了视觉皮层对边缘、线条等基本视觉特征的分层处理机制,该发现后来荣获1981年诺贝尔生理学或医学奖。 ADSFAEQWER353423413434
结构与位置编辑本段
复杂细胞主要分布在后侧大脑的视觉皮层区域,包括: ADFASDFAF23RQ23R
- 初级视皮层(V1):位于枕叶,是视觉信息处理的第一站,复杂细胞在此具有方向选择性。
- 次级视皮层(V2、V3):接收V1输入,参与更复杂的视觉整合。
- Brodmann 19区(V3/V3A):位于枕叶外侧,与物体识别和运动感知密切相关。
这些区域共同构成视觉处理的层级网络,复杂细胞在其中作为中间层的特征检测器,将简单细胞输出的局部边缘信息转化为宏观轮廓。
ADFASDFAF23RQ23R
功能与机制编辑本段
方向选择性与位置不变性
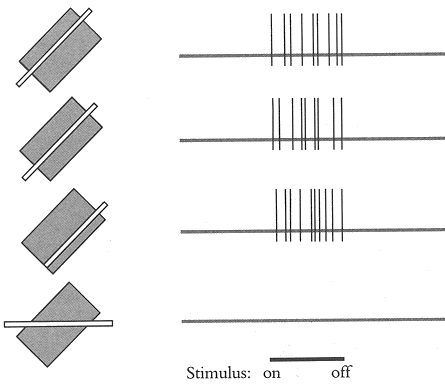
复杂细胞的核心特性是方向选择性,即对某一特定朝向的线条或边缘反应最强,且这种反应在感受野内具有位置不变性。不同于简单细胞需要刺激位于感受野的特定子区域才能激发,复杂细胞可以在整个感受野内对相同朝向的刺激产生均匀反应。例如,一个对水平方向敏感的复杂细胞,无论水平线条出现在感受野的上部、中部还是下部,都会放电。 ADSFAEQWER353423413434
感受野结构
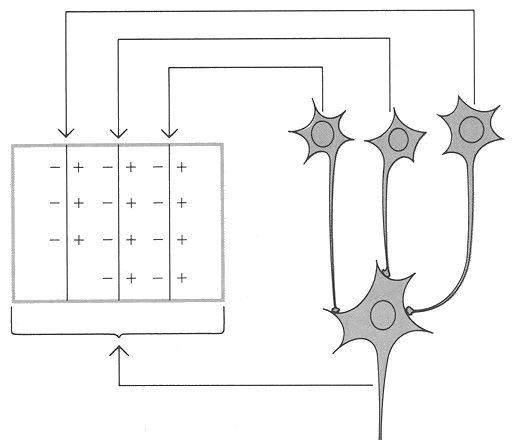
复杂细胞的感受野通常比简单细胞更大,由多个简单细胞的输入汇聚而成。其感受野没有明确的ON区和OFF区,而是对边缘或线条的整体方向作出响应。研究表明,复杂细胞的反应也依赖于亮度的对比度:足够的明暗对比才能激活这些细胞,弱对比度可能无法诱发动作电位。 ADFASDFAF23RQ23R
运动检测
复杂细胞对垂直于其偏好方向的移动刺激表现出强烈反应。例如,偏好垂直方向的复杂细胞会对水平移动的垂直线条产生持续发放。运动方向与偏好方向的关系决定了细胞反应的强弱,这为视觉系统感知物体运动提供了基础。
层级信息处理
视觉信息从视网膜出发,经过外膝体(LGN)到达简单细胞,再由简单细胞汇聚到复杂细胞。简单细胞通过将光感受器(视杆细胞和视锥细胞)传来的亮度对比信息编码为边缘方向,复杂细胞则将这些局部边缘整合成连续的轮廓。这一过程可表示为:
ADSFAEQWER353423413434
简单细胞 → 复杂细胞 → 更高层视觉区域(如V4、IT区),最终实现物体识别。 ADFASDFAF23RQ23R
与简单细胞的比较编辑本段
| 特征 | 简单细胞 | 复杂细胞 |
|---|---|---|
| 感受野结构 | 明确的ON/OFF区域,空间总和性 | 无明确分区,对整体方向反应 |
| 位置依赖性 | 刺激必须位于精确位置 | 位置不变性,大范围感受野 |
| 方向选择性 | 有,但受位置限制 | 强且位置稳定 |
| 主要功能 | 边缘检测(局部) | 轮廓整合与运动感知 |
| 输出目标 | 复杂细胞和其他简单细胞 | 高级视觉区域 |
在视觉通路中的角色编辑本段
复杂细胞在两条主要视觉通路(是什么和在哪里)中发挥关键作用: ADFASDFAF23RQ23R
- 腹侧通路(what通路):经V2、V4至下颞叶皮层,负责物体识别。复杂细胞提供的轮廓信息是形状感知的基础。
- 背侧通路(where通路):经V3、MT至顶叶皮层,负责空间定位和运动分析。复杂细胞的运动检测功能在此通路中至关重要。
例如,在Brodmann 19区(V3),复杂细胞参与区分物体身份(什么)和空间位置(哪里),整合来自简单细胞的边缘信息以决定物体边界。 ADFASDFAF23RQ23R
生物学意义与应用编辑本段
复杂细胞的研究揭示了视觉系统从低级特征到高级对象识别的层级处理机制。这些发现不仅深化了对脑功能的理解,还启发了人工智能中卷积神经网络的设计。例如,深度学习的卷积层模拟了简单细胞和复杂细胞的功能,通过局部感受野和池化操作实现特征不变性。临床上,复杂细胞的损伤可能与视觉失认症或运动感知障碍相关。
总结编辑本段
复杂细胞作为视觉皮层中重要的特征检测神经元,通过方向选择性和位置不变性,将简单细胞的局部边缘信息整合为整体轮廓,支持物体识别和运动感知。其发现对于理解大脑的视觉处理流水线具有里程碑意义,并持续影响神经科学和人工智能的发展。 ADFASDFAF23RQ23R
参考资料编辑本段
- Hubel, D. H., & Wiesel, T. N. (1959). Receptive fields of single neurones in the cat's striate cortex. Journal of Physiology, 148(3), 574-591.
- Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. Journal of Physiology, 160(1), 106-154.
- Movshon, J. A., Thompson, I. D., & Tolhurst, D. J. (1978). Spatial and temporal contrast sensitivity of neurones in areas 17 and 18 of the cat's visual cortex. Journal of Physiology, 283(1), 101-120.
- Livingstone, M. S., & Hubel, D. H. (1988). Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science, 240(4853), 740-749.
- Carandini, M., Demb, J. B., Mante, V., Tolhurst, D. J., Dan, Y., Olshausen, B. A., ... & Rust, N. C. (2005). Do we know what the early visual system does? Journal of Neuroscience, 25(46), 10577-10597.
- Yamins, D. L., & DiCarlo, J. J. (2016). Using goal-driven deep learning models to understand sensory cortex. Nature Neuroscience, 19(3), 356-365.
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。