内含子保留
定义与基本概念
内含子保留是可变剪接的一种特殊形式。在真核生物中,基因通常由外显子(编码区)和内含子(非编码区)交替排列组成。在经典的剪接过程中,所有内含子均被剪接体精确切除,外显子依次连接形成成熟的mRNA。然而,在内含子保留事件中,一个或多个内含子序列被“保留”在最终的成熟转录本中。这些被保留的内含子可能含有提前终止密码子(PTC),导致转录本被NMD途径降解;也可能影响mRNA的出核运输,或产生新的蛋白质亚型。 ADSFAEQWER353423413434
在植物中,内含子保留是最常见的可变剪接事件;在动物中,虽然曾被认为几乎不存在或不重要,但近年来的研究揭示它在动物中同样普遍。
分子机制
2.1 顺式作用元件
内含子保留的发生受到多种顺式作用元件的调控: ADFASDFAF23RQ23R
- 剪接位点强度:被保留的内含子通常具有较弱的剪接位点(特别是5'剪接供体位点和3'剪接受体位点),导致剪接体难以识别并切除这些内含子。
- 内含子长度:短内含子更容易被保留。在人类中,被保留的内含子通常比被切除的内含子更短。
- GC含量:被保留的内含子通常具有较高的GC含量。
- 内含子在基因中的位置:内含子保留倾向性依赖于内含子在基因中的位置,在非翻译区(UTR)中富集。
- 染色质修饰:特定组蛋白修饰(如H3K27ac)与内含子保留相关。
2.2 反式作用因子
多种反式作用因子参与调控内含子保留: ADFASDFAF23RQ23R
- RNA结合蛋白(RBPs):多种RNA结合蛋白可特异性结合内含子或外显子中的调控序列,促进或抑制特定内含子的保留。
- 剪接体:剪接体的组装效率和催化活性直接影响内含子是否被切除。
- 外显子连接复合物(EJC):EJC在剪接后沉积于外显子-外显子连接处,可影响下游内含子的识别。
- 蛋白质精氨酸甲基转移酶(PRMTs):I型和II型PRMTs通过Sm和CHTOP蛋白的甲基化反向调控核内滞留内含子的转录后加工。
2.3 RNA聚合酶II延伸速率
RNA聚合酶II的转录延伸速率是调控内含子保留的重要因素。快速的延伸速率可能使剪接体来不及识别弱剪接位点,从而导致内含子被保留。 ADSFAEQWER353423413434
2.4 核内滞留内含子与“滞留”内含子的区别
研究者将内含子保留进一步细分为两种类型: ADFASDFAF23RQ23R
- “滞留”内含子(Detained introns):指被转录后滞留在细胞核内、等待后续剪接的内含子。这些内含子已与剪接体结合但尚未被切除,可在特定信号下被“释放”并完成剪接。
- “保留”内含子(Retained introns):指最终保留在成熟转录本中、被输出到细胞质的内含子。
这两种类型的区分强调了内含子保留并非总是转录本的“终点”,而可能是一个可逆的调控中间状态。
生物学功能
3.1 基因表达的下调
内含子保留最经典的功能是下调基因表达: ADSFAEQWER353423413434
- NMD途径(IR-NMD):保留的内含子若含有提前终止密码子(PTC),该转录本将被无义介导的mRNA衰变(NMD)途径识别并降解。
- 核内滞留:大多数内含子保留转录本被滞留在细胞核内,无法被翻译。部分核内滞留的IR-RNA可被外泌体降解,也可稳定存在并等待信号触发转录后剪接。
3.2 产生新的蛋白质亚型
部分内含子保留转录本可以被输出到细胞质并翻译。如果保留的内含子不破坏开放阅读框(ORF),或位于5'UTR/3'UTR中,它们可能产生具有新功能的蛋白质亚型。 ADFASDFAF23RQ23R
3.3 应激响应与发育调控
内含子保留在细胞应激响应中发挥“分子开关”的作用。在缺氧、热激、感染等应激条件下,内含子保留模式发生动态变化,帮助细胞快速调整基因表达程序。程序性内含子保留被用于控制多种发育和应激诱导的基因表达程序。内含子保留在神经元、心脏、造血和免疫发育过程中表现出细胞类型和组织特异性的表达模式。
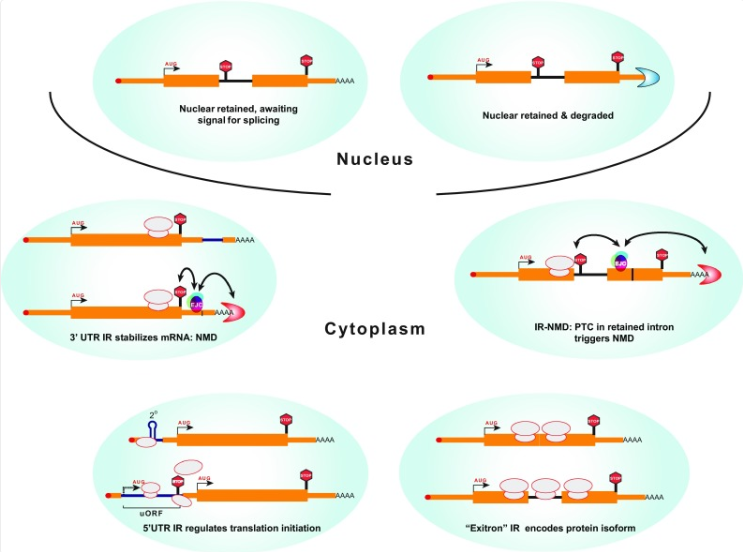 图1 内含子保留的四种主要功能后果
图1 内含子保留的四种主要功能后果
与疾病的关系
内含子保留的异常与多种疾病密切相关: ADSFAEQWER353423413434
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。